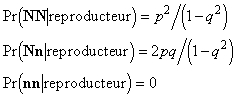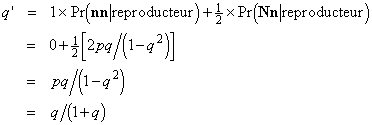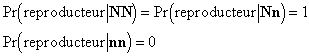
Soit un locus à deux allèles, N et n (nous reprenons les notations relatives au gène de sensibilité à l'halothane chez le porc). L'allèle N, de fréquence p, est dominant ; l'allèle n, de fréquence q, est récessif. Nous nous plaçons dans une population à générations séparées, de taille infinie, où il n'y a ni migration ni mutation. Nous supposons que le régime de reproduction est panmicitique : les trois génotypes, NN, Nn et nn, se trouvent, avant sélection, avec les fréquences respectives p2, 2pq, et q2.
La sélection s'opère de la façon suivante : les individus de phénotype [N] participent tous à la reproduction, les individus de phénotype [n] sont tous éliminés. Nous pouvons alors écrire les probabilités conditionelles pour qu'un individu devienne reproducteur sachant son génotype :
NB. On obtiendrait le même résultat par la suite si, au lieu de 1, on affectait les deux premiers génotypes d'une même probabilité quelconque (mais non nulle, bien sûr).
En combinant les fréquences génotypiques avant sélection et les probabilités conditionnelles de devenir reproducteur, on calcule facilement la probabilité pour qu'un individu de génotype quelconque devienne reproducteur (cette probabilité est égale à la fraction de la population qui se reproduira) :
En appliquant le théorème de Bayes à cette situation, nous calculons la probabilité pour un individu d'être d'un génotype donné, sachant qu'il est reproducteur :
Ces probabilités nous donnent les contributions relatives de chaque génotype au pool de gènes de la génération suivante (on vérifiera que leur somme est bien égale à 1). Nous en déduisons la fréquence q' de l'allèle n à la génération suivante :
Comme q est strictement positif, q' est inférieur à q (si q valait zéro, le problème que nous sommes en train de traiter ne se poserait pas). Ainsi, comme nous pouvions le prévoir, la sélection contre le phénotype correspondant à l'homozygote récessif a pour effet de réduire la fréquence de l'allèle concerné.
Si cette forme de sélection se poursuit, nous pouvons caractériser la vitesse de diminution de la fréquence de l'allèle récessif. Soit qt la fréquence de l'allèle n à la génération t. En reprenant l'expression ci-dessus, on montre que qt s'exprime simplement en fonction de la fréquence initiale de l'allèle (q0) :
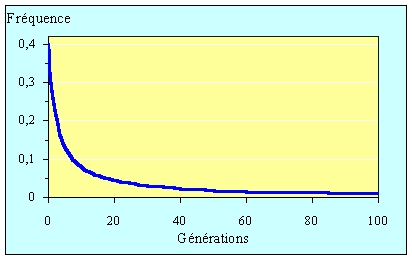
La figure ci-dessous
représente l'évolution de la fréquence d'un allèle récessif sous cette forme de
sélection. On voit qu'à partir d'une valeur moyenne, cette fréquence chute rapidement.
Par contre, dès que cette fréquence est faible, elle diminue très lentement. A partir
de l'équation précédente, on montre facilement que le nombre de générations
nécessaire pour diviser par deux la fréquence de l'allèle récessif est égal à
l'inverse de cette fréquence : il faut 2 générations pour passer de 0,5 à 0,25, mais
il faut 10 générations pour passer de 0,1 à 0,05 ! Ce phénomène s'explique aisément.
En effet, la proportion d'homozygotes parmi les porteurs de l'allèle contre-sélectionné
est d'autant plus forte que sa fréquence est élevée. Lorsque cette fréquence est
faible, la très grande majorité des porteurs de l'allèle sont des hétérozygotes, dont
le phénotype ne donne pas prise à la sélection. On comprend ainsi que, dans divers
types de population, des allèles récessifs induisant des anomalies graves peuvent se
maintenir à de faibles fréquences pendant de nombreuses générations, malgré la
contre-sélection dont ils font l'objet (penser aux nombreuses anomalies à déterminisme
simple chez l'homme ou chez les animaux).
Evolution de la fréquence d'un allèle récessif ayant une fréquence initiale de 0,4 sous l'effet de l'élimination systématique à chaque génération des homozygotes correspondants.
Pour en savoir plus sur la sélection en génétique des populations... (retour au site de l'université de Tours du réseau GENET) |
|
UFR Génétique, élevage et reproduction (AgroParisTech) © gestion des pages - remarques & suggestions : Xavier Rognon (xavier.rognon at agroparistech) - mise à jour : Novembre 1999 |